Le site des ophtalmologistes de France
Espace Encyclopédie
Encyclopédie de la vue

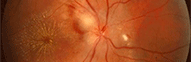




La vision des bovins
Nous remercions Alexandre DIMBERTON. Thèse vétérinaire N° 92, année 1999
Anatomie
L’oeil est constitué d’un organe globuleux, " le bulbe de l’oeil ", logé dans l’orbite. Il est orienté par des muscles et protégé par les paupières dont les mouvements à la surface de l’œil sont facilités par les sécrétions de diverses glandes lacrymales.
Le bulbe de l’œil est délimité par une coque composée de trois tuniques (fibreuse, vasculaire et nerveuse) qui enferme les milieux optiques transparents.
- la tunique fibreuse, externe, comprend :
- la sclère, blanche opaque et solide qui forme les 4/5 postérieurs du bulbe et en assure la rigidité
- la cornée, transparente placée en avant de l’œil. Cette " fenêtre " permet à la lumière de rentrer dans l’œil.
- la tunique vasculaire, intermédiaire se compose de vaisseaux, de nerfs et de tissus élastiques. Elle comprend :
- la choroïde qui correspond à la partie caudale de cette tunique. Elle possède une couche qui reflète la lumière : le tapis clair, facilement identifiable la nuit lorsqu’il renvoie la lumière de phares de voitures par exemple. Il existe chez la plupart des animaux domestiques ou sauvages mais il est absent chez l’homme et le cochon. Le tapis se comporte comme un miroir, il réfléchit la lumière non absorbée par les photorécepteurs (cônes et bâtonnets ) de la rétine qui sont à nouveau stimulés par cette lumière réfléchie. Ce phénomène de réflexion explique en partie la meilleure vision nocturne des animaux par rapport à l’homme.
- le corps ciliaire qui fixe le cristallin et permet l’accommodation. Il secrète l’humeur aqueuse.
- l’iris qui constitue un diaphragme dont l’ouverture, dénommée pupille, varie en fonction de la luminosité, ou de la distance de l’objet fixé. Elle diminue si la lumière est intense, les rayons lumineux pouvant altérer les pigments visuels (phénomène d’éblouissement), elle augmente pour laisser " entrer " plus de lumière si le milieu est sombre. Cette variation réflexe dépend de facteurs musculaires (muscle constricteur) et vasculaires (turgescence des nombreux vaisseaux de l’iris).
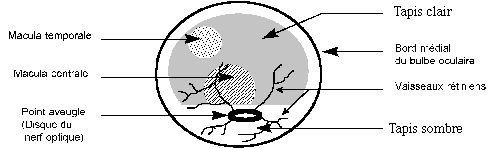
- la tunique nerveuse, la plus interne, encore appelée rétine. Elle se différencie en plusieurs couches (schéma 15):
- une couche non vasculaire et sensorielle : elle comprend les cellules sensorielles ou photorécepteurs, entités élémentaires de la vision. On distingue les bâtonnets qui contiennent des pigments visuels à base de rhodopsine et les cônes dont les pigments à base de photopsine sont sensibles aux différentes longueurs d’ondes de la lumière. Les cônes sont ainsi responsables de la discrimination des couleurs. Ces récepteurs ne sont pas uniformément présents dans la rétine (schéma 16). Ils sont absents au niveau du disque du nerf optique (zone aveugle) et concentrés dans la macula centrale chez l’homme et dans la macula temporale chez les grands ongulés. Grâce à cette concentration, la macula est plus sensible au détail que le reste de la rétine, et permet une visualisation fine des mouvements.
- une couche vasculaire et nerveuse formée de neurones périphériques d’association (cellules bipolaires) et de neurones centraux (cellules ganglionnaires) dont les axones forment le nerf optique.
- une couche non vasculaire et sensorielle : elle comprend les cellules sensorielles ou photorécepteurs, entités élémentaires de la vision. On distingue les bâtonnets qui contiennent des pigments visuels à base de rhodopsine et les cônes dont les pigments à base de photopsine sont sensibles aux différentes longueurs d’ondes de la lumière. Les cônes sont ainsi responsables de la discrimination des couleurs. Ces récepteurs ne sont pas uniformément présents dans la rétine (schéma 16). Ils sont absents au niveau du disque du nerf optique (zone aveugle) et concentrés dans la macula centrale chez l’homme et dans la macula temporale chez les grands ongulés. Grâce à cette concentration, la macula est plus sensible au détail que le reste de la rétine, et permet une visualisation fine des mouvements.

Schéma 15 : Structure de la rétine des bovins (31)
Les milieux optiques transparents comprennent (schéma 18) :
- le cristallin, véritable lentille biconvexe qui concentre et dirige les rayons lumineux sur la rétine. La diminution de la courbure du cristallin secondaire à la contraction du corps ciliaire permet l’accommodation, c’est à dire de focaliser l’image sur la rétine. Une projection en avant ou en arrière de la rétine rendrait l’image flou (phénomène de myopie et d’hypermétropie).
- l’humeur aqueuse en avant du cristallin
- le corps vitré entre le cristallin et la rétine.

Physiologie
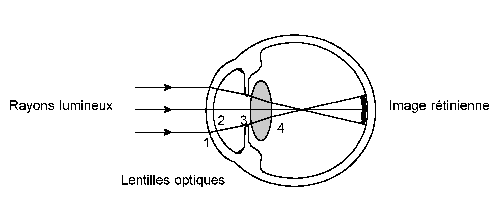
Schéma 19 : Progression des rayons lumineux dans l'œil et formation d'une image rétinienne (76)
Ainsi l’œil comprend un certain nombre de structures qui ont pour rôle de concentrer sur la rétine les rayons lumineux venant de l’extérieur, de façon à donner une image nette et lumineuse. De même, les mouvements des yeux ou ceux de la tête contribuent à placer les images rétiniennes des objets sur la macula afin d’augmenter leur résolution (schéma 19).
Sous l’effet des rayons lumineux, les pigments visuels contenus dans les photorécepteurs se modifient en produisant une dépolarisation de la membrane de l’axone. Ces pigments photolabiles sont constamment régénérés afin d’en maintenir le nombre constant (31).
Le potentiel d’action engendré est transmis à des neurones disposés en couches successives reliées par des synapses : les cellules bipolaires font suite aux récepteurs visuels et entrent en rapport avec les cellules multipolaires ou ganglionnaires dont les axones se réunissent pour constituer le nerf optique. Ces neurones bipolaires et multipolaires condensent les impressions visuelles.
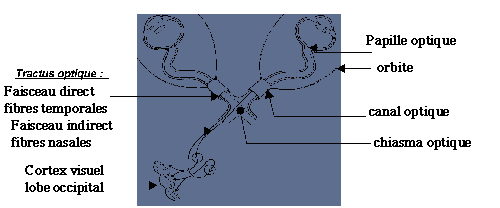
Les deux nerfs optiques s’imbriquent au niveau du chiasma optique puis les fibres nerveuses se dirigent vers la zone visuelle du cortex cérébral (schéma 20).
Plusieurs photorécepteurs, cônes ou bâtonnets, convergent vers une même cellule ganglionnaire. Cette fusion des informations nerveuses détermine la sensibilité définie comme la faculté à détecter un stimulus et l’acuité qui correspond à l’aptitude à percevoir les détails.
Si les cellules ganglionnaires sont peu nombreuses face à une importante population de photorécepteurs, la sensibilité prédomine sur l’acuité. En effet, la cellule ganglionnaire fait la somme des potentiels d’action qu’elle reçoit, ainsi le seuil d’activation est plus rapidement atteint. Par contre, plusieurs détails perçus par des photorécepteurs différents seront "assemblés" pour n’en former plus qu’un, ainsi l’acuité sera diminuée.
A l’inverse, l’acuité visuelle sera favorisée par un plus faible nombre de récepteurs par rapport aux cellules ganglionnaires.
Les bâtonnets convergent proportionnellement aux cônes en plus grand nombre vers une même cellule ganglionnaire, ils sont donc plus sensibles mais ils ne permettent pas une bonne acuité visuelle. Cette sensibilité des bâtonnets est utile dans des conditions de faible luminosité. Inversement en plein jour, la stimulation excessive des cellules ganglionnaires entraîne l’éblouissement (42).
Pour résumer, les cônes interviennent majoritairement dans la vision diurne et dans la distinction des couleurs, les bâtonnets eux jouent un rôle important dans la vision nocturne. Autrement dit, les cônes sont responsables de l’acuité visuelle et chromatique, les bâtonnets de la sensibilité lumineuse.
Après ces rappels anatomo-physiologiques, nous allons nous intéresser plus particulièrement à la vision des bovins et à ces conséquences pratiques pour la contention.
Applications des particularités des bovins à leur manipulation
La vision des bovins n’est pas identique point pour point à celle de l’homme, ses particularités doivent être prises en compte dans la conception des systèmes de contention (42).
L’accommodation
L’accommodation est l’aptitude de l’œil à subir des modifications afin de donner une vision nette d’un objet (mécanisme de mise au point).
Le point le plus proche qui puisse être vu nettement correspond au punctum proximum. Au contraire, la vision des objets éloignés va théoriquement à l’infini, définissant ainsi le punctum remotum.
Comme nous l’avons vu précédemment, le mécanisme de l’accommodation repose sur la déformation du cristallin par le corps ciliaire.
Kendrick (75) a observé que " les moutons possèdent un faible pouvoir d’accommodation ". De plus, d’après Ducks (44), les muscles ciliaires sont peu développés et la capsule du cristallin n’a pas une qualité ni une dimension favorable à l’accommodation.
En réalité, leur faculté d’accommodation est différente de la nôtre. Chez l’homme, l’œil est normalement adapté à la vision à l’infini ; pour la vision rapprochée, il doit mettre en jeu le muscle ciliaire qui tend le cristallin par l’intermédiaire des fibres zonulaires.
A l’inverse, chez les herbivores, le cristallin est globuleux par conséquent les images proches sont naturellement nettes, et les muscles ciliaires se contractent pour " mettre au point " à l’infini (schéma 21). L’accommodation naturelle des éléments proches est adaptée au mode d’alimentation des herbivores qui impose une vision nette de l’herbe proche d’eux pour la brouter. A l’inverse,les carnivores, prédateurs, ont plus intérêt à repérer leurs proies de loin.
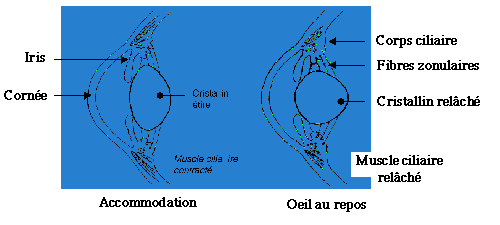
Schéma 21 : Mécanisme de l’accommodation chez les bovins (44)
En conséquence, pour approcher un bovin en partant de loin dans une pâture, le manipulateur doit progresser lentement afin que l’animal ait le temps d’accommoder et de bien identifier ce qui s’approche de lui.
Acuité visuelle et sensibilité
Le rôle de la rétine est toujours un compromis entre la sensibilité et l’acuité visuelles.
Ces deux propriétés de la rétine dépendent de plusieurs facteurs :
- type et densité des photorécepteurs présents (cf. II-B-2.1).
- présence d’un tapis ou non. Le tapis en reflétant la lumière augmente la sensibilité au détriment de l’acuité
- grandeur du champ visuel qui s’accompagne d’une diminution de l’acuité visuelle
- forme du cristallin
- richesse des connexions nerveuses
- aptitude du cerveau à analyser l’information perçue par l’œil.
L’observation et la synthèse de ces différents facteurs permettent de qualifier le système optique d’un animal soit de sensible, c’est à dire, permettant la détection d’un stimulus, soit de bonne acuité, c’est à dire percevant les détails. Néanmoins sensibilité et acuité visuelle convenables peuvent coexister pour un même animal.
Le ratio cônes / bâtonnets des bovins qui possèdent, rappelons-le, une dominante de bâtonnets, ainsi que la présence d’un tapis oriente l’œil des bovins vers une bonne sensibilité mais une acuité visuelle réduite.
Ainsi leur vision est profonde, ils perçoivent une personne à 900 mètres (même sensibilité que l’homme) mais grossière. Ceci explique que les herbivores sauvages ne réagissent pas à la vue d’un homme éloigné et immobile car ils ne peuvent l’identifier.
Entsu, Dohi et Yamada (29) ont étudié l’acuité visuelle des bovins par une expérience basée sur l’apprentissage. Les génisses avaient le choix entre deux auges identifiées par des panneaux différents, l’une contenant du blé, l’autre vide. Le choix entre ces deux auges devaient se faire à une distance déterminée par une cloison centrale. La place des deux auges alternait régulièrement (schéma 22).
Lorsque les génisses choisissaient régulièrement l’auge contenant le blé, l’acuité visuelle était déterminée en augmentant la distance de choix par prolongement de la cloison centrale. L’acuité visuelle était considérée comme dépassée lorsque les génisses choisissaient indifféremment les deux auges. Cette limite fût atteinte lorsque la distance de choix était à 4,3 mètres pour une taille des panneaux de 2 centimètres.
L’acuité visuelle apparaît nettement inférieure à celle de l’homme. La reconnaissance visuelle des bovins est donc réduite, d’autres facteurs comme l’odeur semblent primordiaux.
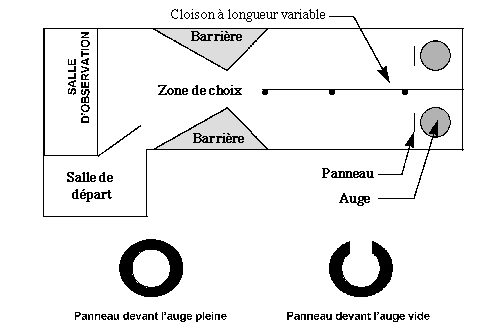
Schéma 22 : Principe de l’expérience de Entsu, Dohi et Yamada (29)
Cette expérience s’intéresse à l’acuité visuelle statique, c’est à dire, au plus petit détail immobile perçu par l’œil. Néanmoins Mc Cormick (29) distingue d’autres types d’acuité visuelle notammentl’acuité dynamique qui correspond à la discrimination des détails lorsque l’objet est en mouvement. L’aire centrale en forme de strie (Area centralis striae formis) de la rétine des bovins leurs permet de déceler des mouvements imperceptibles pour l’homme.
Par exemple, lorsqu’un homme bouge ses bras de façon circulaire, la vache distingue une série de bras décrivant un demi-cercle alors qu’un homme voit le bras seulement au début et à la fin du mouvement. Schématiquement, la vache réalise une série de photos alors que l’homme réalise un film (schéma 23).

Schéma 23 : Perception des mouvements chez l’homme et les bovins
Cette extrême décomposition des mouvements explique la peur des bovins face à des mouvements trop rapides.
En conséquence, lors de l’approche d’un bovin, il faut se déplacer lentement et régulièrement afin de ne pas l’effrayer et immédiatement s’arrêter dès qu’il bouge même si l’on a un pied en l’air. Les mouvements saccadés sont à éviter car ils passent par une phase d’accélération.
A l’inverse pour effrayer des vaches, il faut réaliser des mouvements rapides. Par exemple, pour stopper ou barrer le chemin à une vache, il faut agiter rapidement les bras pour réaliser un demi-cercle. L’emploi d’un bâton qui en prolongeant les bras augmente le diamètre du demi-cercle améliore cette technique.
Remarque : un chien berger fait peur à un troupeau de vaches non par sa taille mais par ses mouvements rapides le long du troupeau. Les Bergers des Pyrénées ont d’ailleurs toujours été sélectionnés sur leur vivacité.
D’autre part, la forte sensibilité visuelle des bovins entraîne des phénomènes d’éblouissements par des lumières d’aspect peu intense pour notre vision. Il peut s’agir de vitres qui reflètent la lumière, d’un pont en inox d’une bétaillère, ou tout simplement des rayons lumineux qui passent à travers une vitre.
On peut observer ce phénomène dans des couloirs de contention éclairés latéralement par des fenêtres (41). Les rayons lumineux pénètrent dans la pièce et forment des carrés lumineux sur le sol. Les bovins ralentissent ou s’arrêtent dès qu’il faut " franchir " un de ces obstacles lumineux.
Tout contraste fait peur aux bovins. Ainsi le changement de couleur d’un sol entre deux pièces peut stopper les animaux.
Par conséquent, il faut manipuler les bovins à l’intérieur d’une pièce uniformément éclairée et dépourvue de reflets lumineux en obstruant les vitres reflétant le soleil. De même, les ponts de bétaillères en inox devraient être peints en noir pour supprimer tout reflet. Enfin, il est préférable de manipuler les bovins avant le lever du soleil, ils sont alors plus calmes et moins éblouis par le rayonnement solaire.
Par contre, si la lumière ne les éblouit pas, les bovins ont toujours tendance à se déplacer vers le milieu le mieux éclairé, ce sont des animaux diurnes.
Cette observation à deux applications :
- pour charger des bovins dans un camion, deux lampes placées au fond de la bétaillère et sur le pont attirent les animaux à l’intérieur à condition qu’elles ne les éblouissent pas. Cette technique marche d’autant mieux que le chargement se fait dans des conditions de faible luminosité, avant le lever du jour par exemple.
- pour sortir les animaux après l’hivernage, une lampe (phare de tracteur par exemple) placée au milieu du prés attire les animaux si on les manipule avant le lever du jour.
- &emdash; La photorégulation
L’œil s’adapte aux différents niveaux d’intensités lumineuses en modulant le diamètre de la pupille et la quantité de pigments visuels présents dans les photorécepteurs.
Lorsque l’animal passe de l’obscurité au jour, la pupille se referme et des pigments visuels sont détruits afin de diminuer la sensibilité de la rétine. Ces phénomènes sont en général rapides et l’adaptation à la lumière ne pose jamais de véritables problèmes.
A l’inverse lorsque l’animal passe du jour à l’obscurité (étable par exemple), la pupille se dilate et des pigments sont synthétisés.
Mais la synthèse de ces pigments est relativement lente surtout celle de la rhodopsine des bâtonnets qui, rappelons-le, intervient principalement dans la vision en milieu sombre. Le temps d’adaptation à l’obscurité varie entre les espèces en fonction de leur capacité de synthèse des pigments visuels. Il est par exemple de 3 minutes pour les bovins contre 30 secondes pour l’homme (facteur de multiplication de 6).
Ainsi pour faire rentrer un bovin dans une étable lorsqu’il vient de l’extérieur, le manipulateur devra attendre environ 3 minutes dès lors que l’animal regarde à l’intérieur. Ce temps paraît excessif, pourtant il permet d’éviter de nombreux échecs.
Le champ de vision binoculaire se définit comme la zone de superposition des deux champs de vision monoculaires de chaque œil.
La position latérale des yeux et la pupille de forme rectangulaire confèrent aux bovins une vision panoramique à dominante monoculaire. Un champ de vision de 330° leurs permet de voir tout ce qui se passe autour d’eux sans même bouger la tête (23). Les herbivores peuvent ainsi détecter plus facilement les prédateurs (44).
Remarque : Chez les carnivores ou les primates, la position frontale des yeux interdit la vision panoramique (tableau 7).
|
Animal |
|
|
|
|
|
|
|
Homme |
|
|
|
Chat |
|
|
|
Chien |
|
|
|
Lapin* |
|
|
|
Cheval* |
|
|
|
Bovin* |
|
|
|
Mouton* |
|
|
|
Chèvre* |
|
|
* Espèces dont les yeux ont des positions latérales
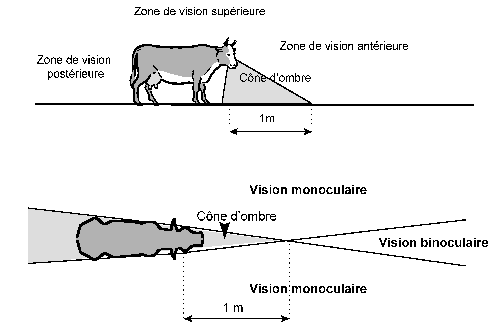
Schéma 24 : Champ de vision panoramique des bovins (44)
Mais cette position latérale des yeux ne présente pas que des avantages. Elle empêche les bovins de concentrer les rayons visuels en deçà d’un point situé à un mètre en avant de l’os frontal. Cette distance varie entre les individus en fonction de la proéminence du front par rapport aux yeux (schéma 24).
Remarque : la taille du cône d’ombre ainsi décrit varie en fonction des auteurs. Bressou l’estime à 15 cm, Rolden à 1 mètre. Néanmoins, nous pouvons l’estimer entre 15 et 20 cm en avant du mufle. Ce cône d’ombre se prolonge plus loin au niveau du sol .
A l’inverse lorsque l’animal a peur ou bien s’énerve, les muscles du bulbe de l’œil se contractent et les yeux se rétractent à l’intérieur des orbites. Cette position encore plus latérale augmente le cône d’ombre (schéma 25). Les bovins deviennent alors très dangereux car ils ne voient plus ce qu’il y a devant eux. Dans tous les cas, il conviendra donc de ne jamais énerver un bovin lorsqu’on le manipule.

Schéma 25 : Modification du champ visuel selon l’état d’excitation de l’animal
Remarque : L’odorat puissant des bovins leur permet d’identifier les objets présents dans ce cône d’ombre, il vient donc compenser cet handicape. Prince (76) observe d’ailleurs que la position latérale des yeux se rencontre chez les animaux qui possèdent un très bon odorat.
La position latérale des yeux réduit également le champ de vision binoculaire, responsable de la perception des distances et de la profondeur relative des objets (notion de relief). Il est estimé compris entre 25° et 50°, les bovins semblent donc mal apprécier les distances. Néanmoins, de multiples expériences (75) tendent à prouver le contraire.
Huston (42) suggère que les bovins, comme beaucoup d’autres espèces (75), utilisent la technique de parallaxe pour estimer une distance. Cette technique basée sur l’angle formé pas deux droites menées de l’objet observé vers deux points d’observation peut être utilisée en vision monoculaire si le bovin bouge son œil ou sa tête. L’apprentissage joue un rôle important (56).
Néanmoins, cette évaluation des distances est moins précise que celle d’une vision binoculaire. En effet, une vache devenue borgne à la suite d’une kératoconjonctivite peut fuir face à un chien situé à 1 mètre comme à 10 mètres d’elle, même s’il l’aborde du côté où elle voit. La notion des distances est fortement perturbée par sa vision monoculaire.
Par conséquent, pour approcher un bovin, nous devons attirer son attention en criant par exemple afin qu’il dirige sa tête vers nous et évalue ainsi correctement la distance qui nous sépare. Cela permet également de ne pas le surprendre.
Avant d’envisager les particularités relatives aux bovins, des rappels sur la notion de couleur sont indispensables.
La lumière blanche est composée d’ondes de différentes longueurs qui correspondent chacune à une couleur donnée.
Les couleurs simples du spectre sont le rouge, l’orange, le jaune, le vert, le bleu et le violet. Elles se composent de couleurs élémentaires :
- le rouge de longueur d’onde comprise entre 647 mm et 723 mm,
- le vert de longueur d’onde comprise entre 492 mm et 575 mm,
- le bleu de longueur d’onde comprise entre 450 mm et 492 mm.
Les objets qui réfléchissent entièrement la lumière sont blancs, ceux qui ne la réfléchissent pas sont noirs. Enfin, les objets colorés ont la propriété d’absorber certaines longueurs d’ondes, celles qu’ils réfléchissent déterminent leur couleur.
Les mécanismes rétiniens de la sensation de couleur ont fait l’objet de nombreuses théories qui, pour la plupart, gravitent autour de la conception trichromique émise par Young en 1807 (44). Selon lui, la sensation de couleur est obtenue par le mélange des trois couleurs élémentaires. Il suffit de supposer l’existence de trois sortes de cônes spécifiquement sensibles aux ondes longues (rouge), moyennes (vert) et courtes (bleu) pour expliquer la sensation colorée obtenue par leur stimulation simultanée.
Etant donné la pauvreté de la rétine des bovins en cônes, eux seuls responsables des impressions chromatiques, nous pouvons concevoir une déficience de la vision des couleurs (76). Néanmoins, comme le souligne Gilbert (33), la simple présence de cônes prouve qu’ils distinguent les couleurs.
Remarque : Certains mammifères qui appartiennent à des espèces nocturnes, ne possèdent pas de cônes, ils ne distinguent donc pas les couleurs. A l’inverse, d’autres possèdent des rétines ne présentant que des cônes (76).
Cette supposition anatomo-histologique est confirmée par diverses expériences basées sur le modèle de Entsu, Dohi et Yamada (schéma 20).
Le panneau positif était une couleur, le panneau négatif un gris de luminosité équivalente. Cette précaution assurait les scientifiques que la réponse des animaux était motivée par la couleur et non la luminosité, c’ est à dire la quantité de lumière réfléchie. Les choix discriminant couleur-gris équivalents permettaient de conclure sur la sensibilité des bovins à une couleur donnée.
Les résultats de toutes les expériences convergèrent. Ils indiquent que les bovins perçoivent et différencient correctement les couleurs de longueur d’onde moyenne et longue (550 mm-700 mm), c’est à dire proches du rouge (77, 69).
Par contre, ils discernent mal les couleurs de faibles longueurs d’onde (400 mm-500 mm), c’est à dire proches du bleu (77, 81, 83, 44). De plus l’orange est souvent confondu avec le jaune (44,83) (tableau 8).
|
|
|
|
|
|
|
|
|
Longueur d’onde (nm) |
|
|
|
|
|
|
|
Pourcentage de discrimination |
|
|
|
|
|
|
On constate donc chez les bovins une certaine faculté à percevoir les couleurs sans que cette propriété semble jouer un rôle bien important dans leur comportement. La vision des couleurs est sans doute inutilisée et les bovins sont plus sensibles à une variation d’intensité lumineuse (44, 85). Kendrick partage cet avis en déclarant (75) " les moutons perçoivent les couleurs mais aucune étude n’a prouvé qu’ils se servaient de cette faculté contrairement à l’utilisation par les bovins des intensités lumineuses ".
Nous avons vu précédemment que les bovins ont peur des reflets lumineux. Ainsi la luminosité de certaines couleurs est un facteur de stress car les rayons solaires réfléchis " éblouissent " en quelque sorte les bovins.
Il s’agit des couleurs lumineuses suivantes :
- les couleurs fluorescentes.
- le blanc. La peur du blanc s’observe lorsque les bovins pénètrent dans une étable récemment blanchie ou dans une salle de traite carrelée en blanc. De même, les vaches ont du mal à traverser les lignes blanches de la voirie, non par crainte des gendarmes mais seulement par peur du blanc. Toute peinture ou vêtement blanc est un facteur de stress pour les bovins, il convient donc de les éviter dans les étables.
- le rouge. Les rubans de chantier rouge et blanc sont régulièrement utilisés lors des descentes d’alpage en Isère. Ils permettent de réaliser des clôtures provisoires de façon rapide et efficace. Les bovins peuvent rester jusqu’à 3 heures dans un enclos ainsi réalisé.
- le jaune et l’orange.
A l’inverse, les couleurs sombres, notamment le noir et le marron, qui réfléchissent peu la lumière rassurent les bovins. Le manipulateur a intérêt à s’habiller dans ces couleurs sombres afin de ne pas stresser les animaux par l’éclat de sa tenue.
La reconnaissance fait intervenir, en plus du sens visuel, le cortex cérébral et les capacités de mémoire.
La reconnaissance de formes et d’individus est bien établie chez les primates. Elle existe chez les bovins mais paraît moins primordiale que la reconnaissance auditive et olfactive.
Toutefois, à leur deuxième montée en alpage, les bovins reconnaissent facilement le trajet. De plus, de nombreuses études basées sur le modèle de l’expérience de Entsu, Dohi et Yamada (schéma 20) en utilisant des panneaux de formes variables démontrèrent que les bovins distinguent et reconnaissent des formes variées (6, 78) (schéma 26).

Schéma 26 : Exemple de formes distinguées par les bovins (75)
Kendrick (57) a mesuré l’activité électrique de la zone visuelle du cerveau d’un mouton lorsqu’il présentait différentes photographies de congénères à l’animal. Il observa que les photographies prises de face stimulaient plus le cerveau que celles prises de profil ou de dos (schéma 27).
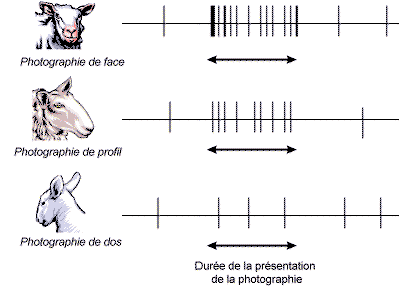
Schéma 27 : Activité électrique du cerveau face à trois photographies de mouton (57)
Ainsi la reconnaissance d’un homme par un bovin passe par la vision de sa face ou de sa silhouette lorsque la distance est trop importante. Par exemple, un homme qui approcherait accroupi un bovin ne serait pas reconnu. Néanmoins ce système est limité et les bovins ne peuvent pas reconnaître seulement par la vue un homme d’un autre. Les stimulations auditives, olfactives interviennent alors.
Le manipulateur doit donc garder une silhouette humaine pour ne pas effrayer les bovins. Une casaque de vêlage qui donne au manipulateur la forme d’un tronc effraye les bovins. Une ceinture placée sur la casaque autour de la taille permet de réduire la peur des animaux et diminue ainsi le risque de recevoir des coups de pied.

![]()
Suivez nous sur...
![]()
SNOF
10 rue Schweighaeuser
CS 40028
67080 STRASBOURG Cedex
Tél. 03 88 35 01 09
Fax. 03 88 25 51 90